| Influence of estrogens on the androgen metabolism in different subunits of human hair follicles |
 |
||
| Antworten | Forum | ||
|
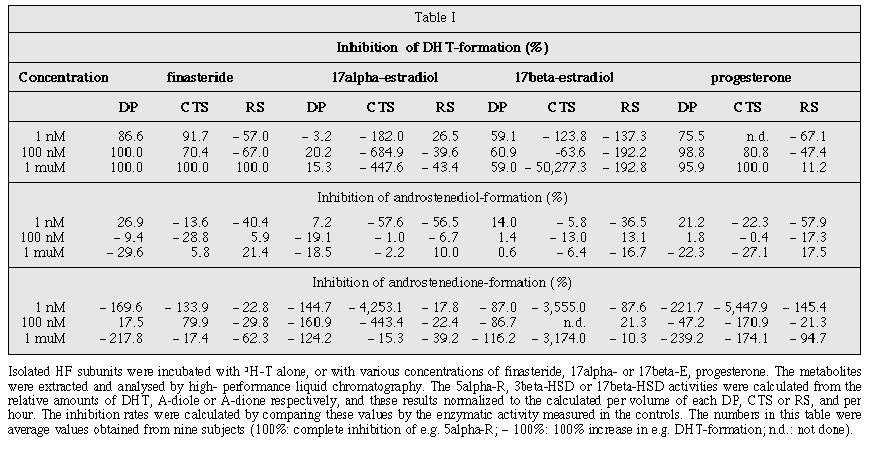
Influence of estrogens on the androgen metabolism in different subunits of human hair follicles Author(s) : S. Niiyama, R. Happle, R. Hoffmann, Department of Dermatology, Philipp University, DeutschhausstraÔe 9, D-35033 Marburg, Germany.. Summary : The molecular pathways involved in estrogen-mediated induction of hair growth in androgenetic alopecia are unknown. Some authors found that estradiol (E) inhibited 5alpha-reductase (5alpha-R) activity and therefore we addressed the question whether 17alpha- or 17beta-E are able to modulate the activity of 5alpha-R, 3beta-hydroxysteroid dehydrogenase (3beta-HSD) or 17beta-hydroxysteroid dehydrogenase (17beta-HSD) in isolated compartments of human hair follicles. For this purpose, scalp biopsies from volunteers were taken and from each biopsy root sheaths, connective tissue sheaths and dermal papillae (DP) were dissected and incubated in the presence of 3H-testosterone (T) and, in addition, either 17alpha-E, 17beta-E, progesterone or finasteride for up to 48 hrs. Thereafter high-performance liquid chromatography analysis of culture supernatants was performed to detect T-metabolites. At the tested concentrations, finasteride was found to be a major inhibitor of dihydrotestosterone (DHT) formation. Even 1 nM finasteride inhibited DHT synthesis in DP by 86% and 1 nM progesterone by 75%. Estrogens were less able to inhibit the synthesis of DHT in DP (e.g. 100 nM 17alpha-E: 20%; 100 nM 17beta-E: 60%). Whether E directly inhibits 5alpha-R in DP's or whether the effect of estrogens might be explained by an increased conversion of T to the weaker androgens such as androstendione (via 17beta-HSD), androstenediol (via 3beta-HSD) or 17beta-E (via aromatase), thereby diminishing the amount of T available for the conversion to DHT, remains to be shown. ARTICLE Androgenetic alopecia (AGA) can be defined as a dihydrotestosterone (DHT)-mediated process, characterized by continuous miniaturization of androgen sensitive hair follicles (HF). The aim of AGA treatment is to reverse or to stop the process of HF miniaturization, which can be accomplished by modifiers of the androgen signal transduction cascade such as 5alpha-reductase (5alpha-R) inhibitors or androgen receptor blockers. Currently available systemic treatment options for AGA therefore involve either the use of antiandrogens such as cyproterone acetate [1] in women, or steroidogenic enzyme inhibitors such as finasteride [2] in men. For the topical treatment of AGA, solutions containing either estradiol benzoate, estradiol valerate, 17beta- or 17alpha estradiol (17beta-E, 17alpha-E) are commercially available in Europe and some studies show an increased anagen and decreased telogen rate after treatment as compared to placebo [3]. At present it is not precisely known how estrogens mediate their beneficial effect on AGA-affected HF. In guinea pigs estrogens have been shown to prolong anagen [3, 4], but in contrast topical application of 17beta-E to the clipped dorsal skin of mice arrested HF in telogen produced a profound and prolonged inhibition of hair growth, while treatment with the biologically inactive stereoisomer, 17alpha-E, had no effect [5-7]. However, some authors found that E might weakly inhibit 5alpha-R activity. Very high doses of estrogens inhibit the testosterone (T) metabolism in rats [8, 9]. Groom et al. [10] reported a direct suppression of 5alpha-R activity in canine prostate explants. Estrogens at very high concentrations (20 çM) have been found to be non-competitive inhibitors of 5alpha-R in granulosa cells from rat ovaries [11]. Other groups did not confirm these data when using genital skin fibroblasts [12]. Moreover, in nearly all experiments showing that estrogens are able to inhibit 5alpha-R in vitro, an inhibition was only present at very high doses of estrogen (e.g. 20 çM) which are usually not applicable in vivo. One study indicated that 17alpha-E at very high concentrations (30 çg/100 mg liver tissues) inhibits 5alpha-R in liver tissues of female rats [13]. The type of 5alpha-R which was inhibited was not determined. The same authors failed to detect a similar effect when using male rat liver tissues. This was attributed to the much smaller total turnover of testosterone in male liver cells. By contrast, the administration of low (physiological) doses of estradiol to rats showed an increase in 5alpha-R activity in prostatic cells [14]. Thus, the use of estrogens for the treatment of AGA is a treatment option which might be able to stop or reverse AGA, but the exact success rates and the underlying mechanisms are unknown or open to question. Therefore we addressed the question whether 17alpha- or 17beta-E are able to modify the androgen metabolic capacity of distinct subunits of human HF in direct comparison with the defined 5alpha-R inhibitors finasteride and progesterone. Materials and methods
Unlabelled dehydroepiandrosterone (DHEA), androstendione (A-dione), androstenediol (A-diol), T and DHT were bought from Sigma (Deisenhofen, Germany). [1,2,6,7-3H(N)]-andost-4-ene-3,17-dione, [1,2,6,7-3H(N)]-andost-4-ene-3,17-diole, [1,2,6,7-3H(N)]-DHEA, [1,2,6,7-3H(N)]-DHT, [1,2,6,7-3H(N)]-T, [1,2,6,7-3H(N)]-androsterone were obtained from NEN Life Science (Boston, MA). Williams E medium was purchased from BioWhittaker/Serva (Heidelberg, Germany) and L-glutamine, insulin, transferrin, sodium selenite and progesterone from Sigma (Deisenhofen, Germany). Finasteride and 17alpha- or 17beta-E were bought from Merck (Rahway, New Jersey). Preparation of HF and their subunits and volume measurement Informed consent was obtained from four female and five male healthy volunteers to perform excisional scalp biopsies from the occiput under local anesthesia. Intact, viable anagen HF were isolated by microdissection as previously described [15]. Single HF were transferred to a petri dish and intact dermal papillae (DP) were microdissected under a stereomicroscope (Fig. 1). The basal tip of the follicle was cut off just above the visible DP, turned inside out, and the DP was then cut off from its capillary stalk. The connective tissue sheath (CTS) and root sheath (RS) were separated by stripping the CTS from the RS by application of gentle pressure with the blunt side of a 26-gauge needle. Virtual volumes were calculated for every DP, CTS or RS immediately after microdissection and shortly before incubation experiments. Digital images were taken at 10 or 40-fold magnification by use of an inverted microscope, a digital camera and the LUCIA M software version 2.995beta (Nikon, Duesseldorf, Germany). The volume of the CTS and the RS were calculated using the "Volume EqCylinder" profile, which is based on a rod model, and the volume of the DP were calculated with the "Volume EqSphere" profile, which is based on a model of the intersection of a ball and a section. The net volume of the CTS was calculated by subtraction of the corresponding RS volume from the CTS gross volume Measurement of enzymatic activity The enzymatic activity was determined with a radiochemical assay. Six DP, three CTS or three RS from one donor were pooled for one measurement and incubated at 37¯ C in a humidified atmosphere containing 5% CO2 for up to 48 hours in supplemented serum-free Williams E medium in the presence of either 50 nM [1,2,6,7-3H]-T (98 Ci/mmol; Amersham, Braunschweig, Germany) alone or in combination with various concentrations (1 muM, 100 nM, 1 nM) of finasteride, 17alpha-E, 17beta-E or progesterone. The reaction products in the medium were extracted twice with three volumes of chloroform/methanol 2:1. Extracts were evaporated at 45¯ C and stored at -70¯ C until analysis. The residue was taken up in 80 mul methanol and aliquots of 20 mul were analysed by high-performance liquid chromatography using a LC-10AD chromatograph (Shimadzu, Tokyo, Japan) and a Beckman Ultrasphere ODS column (5 mum, 4.6 x 250 mm) eluted with acetonitrile/methanol/water 1:3:3. The radioactive metabolites and the remaining substrate were quantified by use of a Flow Scintillation Analyzer 500TR Series (Canberra Packard, Meriden CT). Peaks were identified by their retention times in comparison with 3H-labeled steroids used as standards. The 5alpha-R, 3beta-hydroxysteroid dehydrogenase (3beta-HSD) or 17beta-hydroxysteroid dehydrogenase (17beta-HSD) activities were calculated from the relative amounts of DHT, A-diole or A-dione respectively, and these results normalized to the calculated volume of each DP, CTS or RS as described [15]. The inhibition rates were calculated by comparing these values with the enzymatic activity measured in the controls. Results In our experiments finasteride was the best inhibitor of DHT formation in a dose-dependent manner, followed by progesterone. Even 1 nM finasteride inhibited DHT synthesis in DP by 86% and 1 nM progesterone by 75% in DP, but 1 nM 17alpha-E did not inhibit DHT formation and 1 nM 17beta-E by 59% (Table I). At 100 nM 17alpha-E inhibited DHT formation in DP by 20%, whereas 100 nM finasteride showed a 100% inhibiton. Furthermore we noticed a marked shift to the T metabolites A-diole and A-dione in those experiments where we incubated the CTS with T and estrogens, thus indicating increased 17beta- and 3beta-HSD enzymatic activity. Discussion The local conversion of T to the more potent androgen DHT by 5alpha-R plays a pivotal role in the development of AGA [16], and therefore 5alpha-R has generated great interest as a therapeutic target. The first agent used for this purpose was finasteride, which is an orally active type 2 5alpha-R inhibitor. Clinical studies conducted to treat benign prostatic hyperplasia established the safety profile of this drug [17, 18]. Subsequently oral finasteride was shown to reverse balding in the stump-tailed macaque [19, 20], and dose-finding studies were conducted to compare the efficacy of 5 mg versus 1 mg, 0.2 mg and 0.01 mg finasteride to treat AGA in men, indicating that 1 mg finasteride daily can be regarded as a safe and efficient treatment [21]. Although systemic treatment is clinically effective, topical treatment would be preferred by some patients. For this reason attempts have been made to assess the efficacy of topical finasteride in AGA. The results in humans are disappointing, at present. Topically applied finasteride reduces DHT levels but does not prevent AGA in men [22]. Moreover, both oral and topical finasteride, when applied in pregnant women, implies the risk of feminized male fetuses. In Europe estrogens are used for the treatment of AGA not bearing this contraindication. However, at present the mode of action of estrogens on AGA is unclear. Our results show that estrogens are able to modify the androgen metabolism of distinct subunits of the HF such as the DP, thus diminishing the amount of DHT formed after incubation with T. Whether these effects are directly mediated by the inhibition of 5alpha-R within the HF or are of an indirect nature through estrogen-induced increased conversion of T to the weaker androgens A-diole (via 3beta-HSD), A-dione (via 17beta-HSD) or 17beta-E (via aromatase) thereby diminishing the amount of T available for the conversion to DHT, for example, is not known. Our results indicate that such mechanisms may occur because we noticed an increased concentration of A-diole and A-dione after incubation with estrogens. 17beta-HSD enzyme activity in CTS increased considerably when 17alpha- or 17beta-E was used as compared to finasteride. Whether such mechanisms take place in vivo is not known and should be investigated in the future. It is so far not known whether topically applied E are able to target the DP in vivo at sufficient concentrations. On the contrary finasteride efficacy is well documented in vivo and ex vivo. Finasteride is the only drug where systemic effects in vivo have been correlated with local enzyme inhibition in vivo [23, 24] and also in vitro with fresh scalp evaluation. Therefore, more studies should be performed to evaluate the effects on human hair growth of topically applied estrogen. CONCLUSION Acknowledgements The excellent technical assistance of Elke Wenzel and Andrea Huth (Department of Dermatology, Philipp University) is greatly appreciated. Article accepted on 12/2/01 REFERENCES 1. Ekoe JM, Burckhardt P, Ruedi B. Treatment of hirsutism, acne and alopecia with cyproterone acetate. Dermatologica 1980; 160: 398-404. 2. Hoffmann R, Happle R. Finasteride is the main inhibitor of 5alpha-reductase activity in microdissected dermal papillae of human hair follicles. Arch Dermatol Res 1999; 291: 100-3. 3. Orfanos CE, Vogels L. Lokaltherapie der Alopecia androgenetica mit 17alpha-østradiol: eine Kontrollierte, randomisierte D—ppelblindstudie. Dermatologica 1980; 161: 124-32. 4. Jackson D, Ebling FJ. The activity of hair follicles and their response to oestradiol in the guinea pig Cavia porcellus L. J Anat 1972; 111: 303-16. 5. Oh HS, Smart RC. An estrogen receptor pathway regulates the telogen-anagen hair follicle transition and influences epidermal cell proliferation. Proc Natl Acad Sci USA 1996; 93: 12525-30. 6. Smart RC, Oh HS. On the effect of estrogen receptor agonists and antagonists on the mouse hair follicle cycle. J Invest Dermatol 1998; 111: 175. 7. Stenn KS, Paus R, Filippi M. On the effect of estrogen receptor agonists and antagonists on the mouse hair follicle cycle (reply). J Invest Dermatol 1998; 111: 175. 8. Famsworth WE. A direct effect of estrogens on prostatic metabolism of testosterone. Invest Urol 1969; 6: 423-7. 9. Jenkins JS, McCaffery VM. Effect of oestradiol-17beta and progesterone on the metabolism of testosterone by human prostatic tissue. J Endocrinol 1974; 63: 517-26. 10. Groom M, Harper ME, Fahmy AR, Pierrepoint CG, Griffiths K. The effect of oestrogen on the prostatic metabolism of testosterone in tissue culture. Biochem J 1971; 122: 125-6. 11. Payne DW, Packman JN, Adashi EY. Follicle-stimulating hormone inhibits granulosa cell 5alpha-reductase activity: possible role of 5alpha-reductase as a steroidogenic pubertal switch. J Biol Chem 1992; 267: 13348-55. 12. Beckmann MW, Wieacker P, Dereser MM, Flecken U, Breckwoldt M. Influence of steroid hormones on 5alpha-reductase activity in female and male genital skin fibroblasts in culture. Acta Endocrinol 1993; 128: 161-7. 13. Schriefers H, Wright MC, Rozman T, Hevert F. Hemmung des Testosteron-Stoffwechseis durch 17alpha-Estradiol in Rattenleberschnitten. Arzneimittelforschung 1991; 41: 1186-9. 14. Makeia S, Santti R, Martikainen P, Nienstedt W, Paranko J. The influence of steroidal and nonsteroidal estrogens on the 5alpha-reduction of testosterone by the ventral prostate of the rat. J Steroid Biochem 1990; 35: 249-56. 15. Eicheler W, Happle R, Hoffmann R. 5alpha-reductase activity in the human hair follicle concentrates in the dermal papilla. Arch Dermatol Res 1998; 290: 126-32. 16. Hoffmann R, Happle R. Current understanding of androgenetic alopecia. Part I: Etiopathogenesis. Eur J Dermatol 2000; 10: 319-26. 17. Gormley GJ, Stoner E, Rittmaster RS, Gregg H, Thompson DL, Lasseter KC, Vlasses PH, Stein EA. Effects of finasteride (MK-906), a 5alpha-reductase inhibitor, on circulating androgens in male volunteers. J Clin Endocrinol Metab 1990; 70: 1136-41. 18. Gormley GJ, Stoner E, Bruskewitz RC, Imperato-McGinley J, Walsh PC, McConnell JD, Andriole GL, Geller J, Bracken BR, Tenover JS, Darracott VE, Pappas F, Taylor A, Binkowitz B, Ng J. The effect of finasteride in men with benign prostatic hyperplasia. N Engl J Med 1992; 327: 1185-91. 19. Rhodes L, Harper J, Uno H, Gaito G, Audette-Arruda J, Kurata S, Berman C, Primka R, Pikounis B. The effects of finasteride (Proscar) on hair growth, hair cycle stage, and serum testosterone and dihydrotestosterone in adult male and female stumptail macaques (Macaca arctoides). J Clin Endocrinol Metab 1994; 79: 991-6. 20. Diani AR, Mulholland MJ, Shull KL, Kubicek MF, Johnson GA, Schostarez HJ, Brunden MN, Buhl AE. Hair growth effects of oral administration of finasteride, a steroid 5alpha-reductase inhibitor, alone and in combination with topical minoxidil in the balding stumptail macaque. J Clin Endocrinol Metab 1992; 74: 345-50. 21. Kaufman KD. Clinical studies on the effects of oral finasteride, a type 2 5alpha-reductase inhibitor, on scalp hair in men with male pattern baldness. In: Van Neste D, Randall V, eds. Hair Research for the Next Millenium. Amsterdam: Elsevier, 1996: 363-5. 22. Rushton DH, Norris MJ, Ramsay ID. Topical 0.05% finasteride significantly reduces serum DHT concentrations, but had no effect in preventing the expression of genetic hair loss in men. In: Van Neste D, Randall V, eds. Hair Research for the Next Millenium. Amsterdam: Elsevier, 1996: 359-62. 23. Drake L, Hordinsky M, Fiedler V, Swinehart J, Unger WP, Cotterill PC, Thiboutot DM, Lowe N, Jacobson C, Whiting D, Stieglitz S, Kraus SJ, Griffin EI, Weiss D, Carrington P, Gencheff C, Cole GW, Pariser DM, Epstein ES, Tanaka W, Dallob A, Vandormael K, Geissler L, Waldstreicher J. The effects of finasteride on scalp skin and serum androgen levels in men with androgenetic alopecia. J Am Acad Dermatol 1999; 41: 550-4. 24. Robert JL, Fiedler V, Imperato-McGinley J, Whiting D, Olsen E, Shupack J, Stough D, DeVillez R, Rietschel R, Savin R, Bergfeld W, Swinehart J, Funicella T, Hordinsky M, Lowe N, Katz I, Lucky A, Drake L, Price VH, Weiss D, Whitmore E, Millikan L, Muller S, Gencheff C, Carrington P, Binkowitz B, Kotey P, He W, Bruno K, Jacobsen C, Terranella L, Gormley GJ, Kaufman KD. Clinical dose ranging studies with finasteride, a type 2 5alpha-reductase inhibitor, in men with male pattern hair loss. J Am Acad Dermatol 1999; 41: 555-63.
|
verfasst von: Kai Û 06/04/2004, 18:04:55 ándern |
|
|
Antworten | Spam Alarm |
|
Alle Beitrðge Zum Forum |
|
|


